 Zooplankton of the Great Lakes
Zooplankton of the Great Lakes
Daphnia pulicaria
Taxonomy
Kingdom-
Animalia
Phylum- Arthropoda
Subphylum- Crustacea
Class- Branchiopoda
Order- Cladocera
Family- Daphniidae
Genus- Daphnia
Species- pulicaria
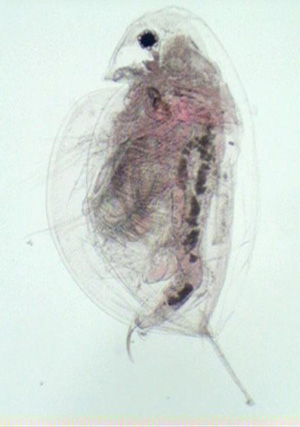
Figure 1. Adult female D. pulicaria with the postabdominal claw extending beyond the carapcae.
Classification
Daphnia pulicaria is very similar to other Daphnia species and is often confused with Daphnia pulex. This similarity has led scientists in the past to place these species in a generalized group called the “Daphnia pulex complex”. Close observation of anatomical and ecological characteristics, however, allows researchers to distinguish D. pulicaria from other species in the genus Daphnia (Brandlova et al. 1972).
Anatomy
Daphnia pulicaria is identifiable by its rounded carapace, large second antennae, pronounced rostrum, and moderately long tail spine (Figure 1). The head of the adult is rather short, rounded, and gently curving near the rostrum (Figure 2). The postabdominal claw has five teeth in the middle pectin that are longer than the distal and proximal pectin (Figure 3) (Urabe et al. 2003). Daphnia pulicaria is distinguishable from other Daphnia species in that it has an ocellus and does not have a pronounced helmet (Brandlova et al. 1972).
Several morphological characteristics distinguish D. pulicaria from the closely related D. pulex. The tail spine of D. pulicaria is often longer than that of D. pulex, although this may be difficult to distinguish. Adult D. pulicaria have a pattern of 6 to 8 rows of elongate polygons between the rostrum and the ocellus. Daphnia pulex also has this pattern but the polygons of D. pulicaria are much longer than wide whereas those of D. pulex are about equal in length and width. Additionally, male D. pulicaria have a short second dorsal abdominal process while male D. pulex have a long second dorsal abdominal process (Brandlova et al. 1972).
Distribution
Daphnia pulicaria is generally found in deeper waters of lakes with highest abundances occurring in the spring (Brandlova et al. 1972, Stich and Maier 2007). Juvenile and adult D. pulicaria distribute differently in lakes based on the quality and availability of food and the presence of predators (Reichwaldt and Abrusán 2007, Reichwaldt 2008). Juveniles are often found in the warmer temperatures of the epilimnion to maximize growth where food is abundant (Reichwaldt 2008). Adults, however, are more visible to planktivorous fish and are therefore usually found in the hypolimnion during the day and exhibit vertical migration at night to feed when the risk of fish predation is lower (Cerny and Bytel 1991, Leibold 1991). Adult D. pulicaria have also been observed migrating to shallow water at night in order to avoid predation from the phantom midge Chaoborus (Gonzales and Tessier 1997).
Feeding Ecology
Daphnia pulicaria primarily consumes algae from the water by filter feeding. The thoracic legs move water which in turn brings food particles towards the mouth. The postabdominal claw is used to clear any debris that becomes lodged near the mouth and feeding appendages. As the amount of debris (filamentous algae) increases, individuals have to spend more time cleaning with the postabdominal claw and less time collecting and ingesting food particles (Gliwicz and Lampert 1990). Daphnia pulicaria has the ability to adapt to this problem. The area and mesh size of the setae (Figure 4) on the feeding appendages (3rd and 4th limbs: Figure 5) increases in response to increased filamentous algae and reduced food quantity. This maximizes feeding efficiency in oligotrophic conditions where food can be scarce and in eutrophic conditions where filamentous algae can dominate (Ghadouani and Pinel-Alloul 2002).
Reproduction
Daphnia reproduce by obligate parthenogenesis where the majority of reproduction is asexual and occurs during favorable conditions. A switch to sexual reproduction occurs when conditions deteriorate in the fall or during periods of crowding. When this occurs, males are produced and copulate with females, producing resting eggs. Resting eggs can tolerate harsh periods including drought and freezing until more favorable conditions for hatching exist (Chen and Felt 1996, Brewer 1998).
Daphnia pulicaria employs specific techniques when finding mates during sexual reproduction. Adults are unable to attract or locate mates using chemical signals but do employ unique swimming patterns. The females swim at slower speeds and move vertically while the males swim quickly in a horizontal direction, making many sharp turns. This behavior increases the amount of contact between males and females. Once contact is made, individuals likely recognize the opposite sex through chemical cues, as the length of contact between males and females is over eight times as long as with the same sex (Brewer 1998). Once a female has been contacted, males are able to select which females they reproduce with. Males are able to detect and avoid copulation with asexually reproducing females and females with resting eggs, thereby only copulating with receptive females (Forró 1997).
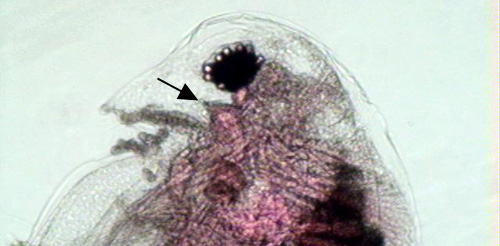
Figure 2. A rounded head lacking a distinct helmet, a pronounced rostrum, and an ocellus (indicated by the arrow) are features of the Daphnia pulex complex.
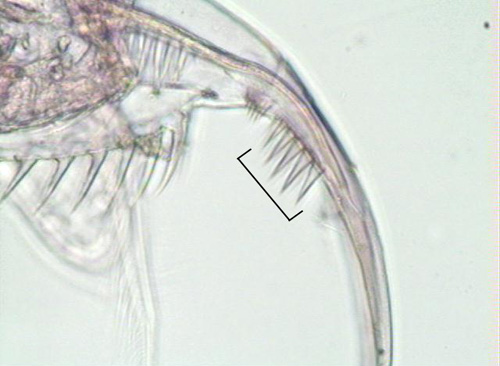
Figure 3. Magnified view of the postabdominal claw of D. pulicaria with its characteristic five long middle pectin.
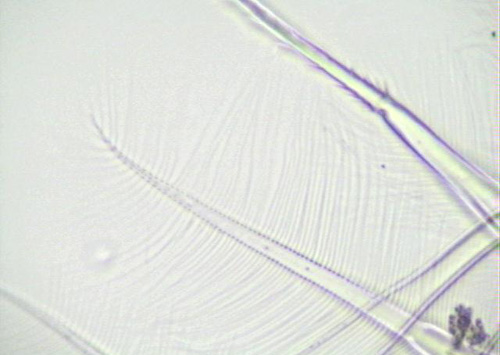
Figure 4. Setae of the third appendage of D. pulicaria. The distance between the setae can be altered in response to the quality and quantity of food available.

Figure 5. The third appendage of an adult D. pulicaria that is used during feeding to filter particles from the water.
Works Cited
Brandlova, J., Z. Brandl, and C. H. Fernando. 1972. The Cladocera of Ontario with remarks on some species and distribution. Canadian Journal of Zoology 50:1373-1403.
Brewer, M. C. 1998. Mating behaviours of Daphnia pulicaria, a cyclic parthenogen: comparisons with copepods. Philosophical Transactions of the Royal Society of London 353:805-815.
Cerny, M., and J. Bytel. 1991. Density and size distribution of Daphnia pulicaria at different fish predation levels. Hydrobiologia 225:199-208.
Chen, C. Y., and C. L. Felt. 1996. Consequences of fall warming for zooplankton overwintering success. Limnology and Oceanography 41:1077-1086.
Forró, L. 1997. Mating behaviour in Moina brachiata (Jurine, 1820) (Crustacea, Anomopoda). Hydrobiologia 360:153-159.
Ghadouani, A., and B. Pinel-Alloul. 2002. Phenotypic plasticity in Daphnia pulicaria as an adaptation to high biomass of colonial and filamentous cyanobacteria: experimental evidence. Journal of Plankton Research 24:1047-1056.
Gliwicz, Z. M., and W. Lampert. 1990. Food thresholds in daphnia species in the absence and presence of blue-green filaments. Ecology 71:691-702.
Gonzalez, M., and A. Tessier. 1997. Habitat segregation and interactive effects of multiple predators on a prey assemblage. Freshwater Biology 38:179-191.
Leibold, M. A. 1991. Trophic interactions and habitat segregation between competing Daphnia species. Oecologia 86:510-520.
Reichwaldt, E. S. 2008. Food quality influences habitat selection in Daphnia. Freshwater Biology 53:872-883.
Reichwaldt, E. S., and G. Abrusán . 2007. Influence of food quality on depth selection of Daphnia pulicaria. Journal of Plankton Research 29:839-849.
Stich, H. B., and G. Maier. 2007. Distribution and abundance of Daphnia pulicaria, a large Daphnia of the “pulex group”, in Lake Constance (Lower Lake). Limnologica 37:303-310.
Urabe, J., S. Ishida, M. Nishimoto, and L. J. Weider. 2003. Daphnia pulicaria, a zooplankton species that suddenly appeared in 1999 in the offshore zone of Lake Biwa. Limnology 4:35-41.